
琥珀校正基因(Ambersuppressors),能够消除琥珀突变所致的表型效应的突变基因。琥珀抑制子突变通常位于一个与琥珀突变无关的基因中,该基因编码一个有突变的反密码子的tRNA,从而当发生琥珀突变产生UAG终止密码子时,仍可将UAG改读为某种氨基酸,使可能被中止的肽链得以继续合成。故又称琥珀抑制子。如突变酪氨酸-tRNA的反密码子3’AUC可识别5’UAG,使酪氨酸插入终止密码,而使多肽链继续合成下去。
精选百科
本文由作者推荐
琥珀校正基因
暂无
琥珀校正基因相关的文章

蚵仔煎是一道将蚵仔、蛋、葱等食材放入加水后的番薯粉浆中一同煎成的饼状物,又叫海蛎煎、蚝烙。蚵仔煎是中国闽南、台湾、潮汕等地区的经典小吃,蚵仔的学名为牡蛎,广东人通常称“牡蛎”为“蚝”,而闽南、东南亚和台湾等地区的闽南人一般称其为“蚵仔”。

潮汕美食节1988年创办于汕头的旅游活动汕头市潮汕美食节于1988年创办,成为汕头市的美食盛事,是我省举办最早,历史最长的旅游活动之一。它对弘扬潮汕饮食文化,拉动内需,刺激消费,促进汕头社会经济和旅游业的发展,打响汕头市“海滨邹鲁,美食之乡”的旅游品牌起到了巨大的推动作用。

汕头金海湾大酒店1991年开业的五星级酒店全国星级饭店五十佳汕头是中国重要的港口城市,五大经济特区之一,著名侨乡,也是著名的海滨城市和百载商埠,素有“粤东门户,海滨邹鲁、美食之乡”之美称,金海湾大酒店是粤东首家五星级商务酒店,位于汕头市中心迎宾广场东南面,坐落于汕头经济特区最繁华的商业金融黄金地带,

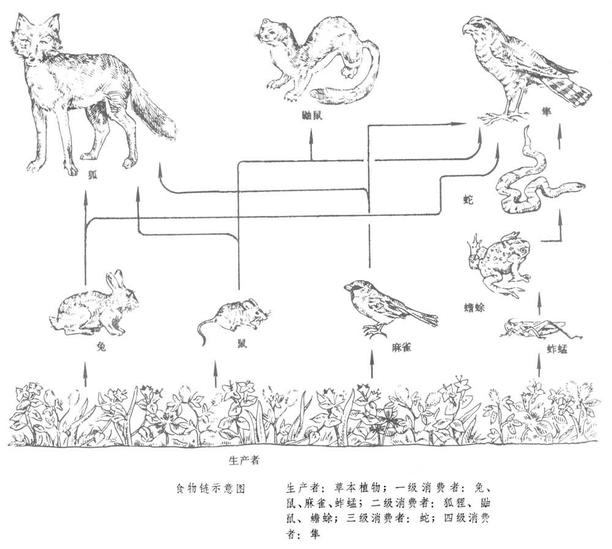
寄生食物链生态系统中贮存于有机物中的化学能在生态系统中层层传导,通俗地讲,是各种生物通过一系列吃与被吃的关系,把这种生物与那种生物紧密地联系起来,这种生物之间以食物营养关系彼此联系起来的序列,在生态学上被称为食物链。按照生物与生物之间的关系可将食物链分为捕食食物链、腐食食物链(碎食食物链)、和寄生食

迪罗乌斯《美食的俘虏》中的种族迪罗乌斯,虚拟角色,是动漫《美食的俘虏》中的一个种族,系远古时代便存在的翼龙类生物,漫画设定为存在过的最强大的龙,此龙一生只长一颗长牙并靠这颗牙成为美食界不可随意侵犯的霸主,此种族最强者为八王之一与美食界第一大陆的统治者。龙王身高4000米,重达8000万吨。

石塘百科
这家伙太懒了,什么都没写!
作者
